NPB 163/PSC 128
Linear receptive field models
Retina
- Since the physiological recordings of retinal ganglion cells
by Hartline, Barlow, and Kuffler during the 1950’s, it has been well known
that these neurons signal the spatial differences in light intensity falling
upon the retina. This is accomplished by the so-called “center-surround”
organization of the receptive field, in which its excitatory and inhibitory
subfields are organized into circularly symmetric regions.
- The shapes of these receptive fields have been modeled
by at least two different types of functions. One is the difference-of-Gaussians
(DOG) function and the other is the Laplacian-of-Gaussian (LOG) function.
The DOG model simply uses the difference of two 2D Gaussians to model the
receptive field shape. The LOG model was proposed by Marr and Hildreth and
uses the second spatial-derivative (hence the term Laplacian) of a Gaussian
to model the receptive field shape. Both of these functions capture reasonably
well the “Mexican hat” shape of retinal ganglion cell receptive fields.
- We can think of the receptive field shape of a retinal
ganglion cell as the linear spatial weighting function of the cell. That
is, we can model the retinal ganglion cell as a linear neuron, where the
receptive field tells us what the weights are. Using the function
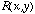 to characterize the receptive field shape using either the DOG or LOG model,
we compute the output of a model retinal ganglion cell as
to characterize the receptive field shape using either the DOG or LOG model,
we compute the output of a model retinal ganglion cell as
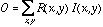
where 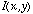 is the input image.
is the input image.
- For a whole array of retinal ganglion cells with identical
receptive fields, we compute the output of each cell in the array as
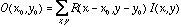
where 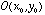 is the output of the retinal ganglion cell whose receptive field is centered
at position
is the output of the retinal ganglion cell whose receptive field is centered
at position 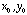 .
.
- Of course, the real situation in the retina is much more
complicated, because the transformation from pixels to retinal ganglion cell
outputs is mediated by many other neurons and complex synapses. But the
above equations nevertheless provide a good first-order approximation of
the function of these cells.
- There are two major varieties of center-surround receptive
fields, on-center/off-surround and off-center/on-surround,
depending on whether the central region is excitatory or inhibitory, respectively.
It is widely thought that the reason for having these two different varieties
is so that both positive and negative changes in intensity can be signaled
with positive-only quantities (action potentials).
- The reason why the retina would compute such a function
in the first place has been the subject of many different theories. One
theory is that it provides a more efficient representation of the image because
it eliminates redundancies due to the similarities of neighboring pixel values
inherent in natural images. Another theory is that it signals the locations
of edges, which is the first step in form or shape analysis.
Cortex
- Proceeding further up the visual pathway, one finds cells
in the visual cortex (area V1) that are orientation-selective, meaning
that they respond to spatial intensity changes only along a certain orientation.
Such cells were named simple-cells and complex-cells by Hubel
& Wiesel, who first described them in the early 1960’s (work for which
they later won the Nobel prize). Here we focus on modeling the structure
of simple-cells.
- Marcelja and Daugman (ca. 1980) have pointed out that the
receptive fields of simple-cells are well-described with a Gabor function,
which is simply a Gaussian modulated sinusoid. In one-dimension, this is
expressed as
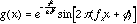
where 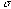 denotes the width of the Gaussian envelope,
denotes the width of the Gaussian envelope, 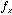 denotes the carrier frequency of the sinusoid, and
denotes the carrier frequency of the sinusoid, and 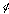 its phase. The larger we make
its phase. The larger we make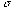 for a fixed frequency
for a fixed frequency 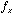 , the more wobbles the function will have. In two-dimensions we have
, the more wobbles the function will have. In two-dimensions we have
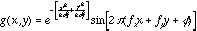
where 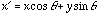 and
and 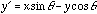 , and
, and 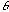 is the orientation of the Gaussian envelope. Note that
is the orientation of the Gaussian envelope. Note that 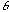 is usually chosen to be the same as the orientation of the sinusoidal carrier,
is usually chosen to be the same as the orientation of the sinusoidal carrier,
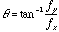 .
.
- Thus, we could think of simple-cells in the visual cortex
as representing the result of computing the inner-product between a Gabor
function and the image at each position in the image, for each orientation
(
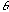 ) and spatial-frequencie (
) and spatial-frequencie (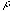 ):
):
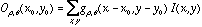
where 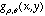 denotes a Gabor function of spatial-frequency
denotes a Gabor function of spatial-frequency 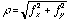 and orientation
and orientation 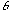 as defined above. Of course the neural images
as defined above. Of course the neural images 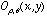 are all intertwined with each other over the cortex, preserving topography
globally and grouping into orientation columns locally.
are all intertwined with each other over the cortex, preserving topography
globally and grouping into orientation columns locally.
- The Gabor function is so-named because it was first discussed
by Denis Gabor, a communication theorist, in the 1940’s (no relation to Zsa-Zsa).
These filters are currently used in image processing, and form useful representations
for compression as well as for image analysis and recognition. One
reason they are thought to be useful (for the brain) is that they produce
a sparse representations of natural images - i.e., only a few neurons need
be active in order to fully represent a natural scene.